Generally, spinal nerves contain afferent axons from sensory receptors in the periphery, such as from the skin, mixed with efferent axons travelling to the muscles or other effector organs. As the spinal nerve nears the spinal cord, it splits into dorsal and ventral roots. The dorsal root contains only the axons of sensory neurons, whereas the ventral roots contain only the axons of the motor neurons. Some of the branches will synapse with local neurons in the dorsal root ganglion, posterior (dorsal) horn, or even the anterior (ventral) horn, at the level of the spinal cord where they enter. Other branches will travel a short distance up or down the spine to interact with neurons at other levels of the spinal cord. A branch may also turn into the posterior (dorsal) column of the white matter to connect with the brain. For the sake of convenience, we will use the terms ventral and dorsal in reference to structures within the spinal cord that are part of these pathways. This will help to underscore the relationships between the different components. Typically, spinal nerve systems that connect to the brain are contralateral, in that the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain.
Cranial nerves convey specific sensory information from the head and neck directly to the brain. For sensations below the neck, the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain. Whereas spinal information is contralateral, cranial nerve systems are mostly ipsilateral, meaning that a cranial nerve on the right side of the head is connected to the right side of the brain. Some cranial nerves contain only sensory axons, such as the olfactory, optic, and vestibulocochlear nerves. Other cranial nerves contain both sensory and motor axons, including the trigeminal, facial, glossopharyngeal, and vagus nerves (however, the vagus nerve is not associated with the somatic nervous system). The general senses of somatosensation for the face travel through the trigeminal system.
Specific regions of the CNS coordinate different somatic processes using sensory inputs and motor outputs of peripheral nerves. A simple case is a reflex caused by a synapse between a dorsal sensory neuron axon and a motor neuron in the ventral horn. More complex arrangements are possible to integrate peripheral sensory information with higher processes. The important regions of the CNS that play a role in somatic processes can be separated into the spinal cord brain stem, diencephalon, cerebral cortex, and subcortical structures.
A sensory pathway that carries peripheral sensations to the brain is referred to as an ascending pathway, or ascending tract. The various sensory modalities each follow specific pathways through the CNS. Tactile and other somatosensory stimuli activate receptors in the skin, muscles, tendons, and joints throughout the entire body. However, the somatosensory pathways are divided into two separate systems on the basis of the location of the receptor neurons. Somatosensory stimuli from below the neck pass along the sensory pathways of the spinal cord, whereas somatosensory stimuli from the head and neck travel through the cranial nerves—specifically, the trigeminal system.
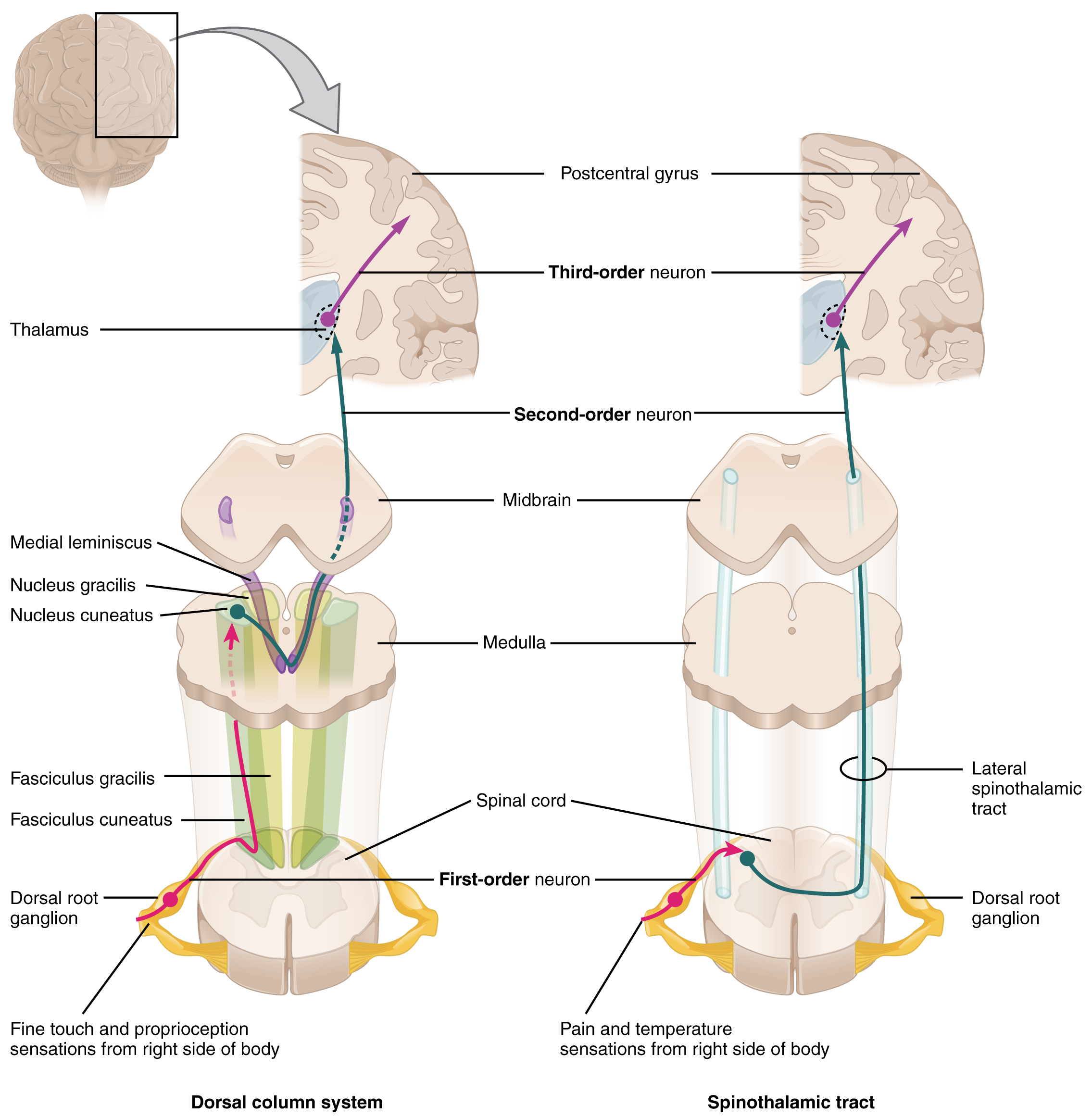
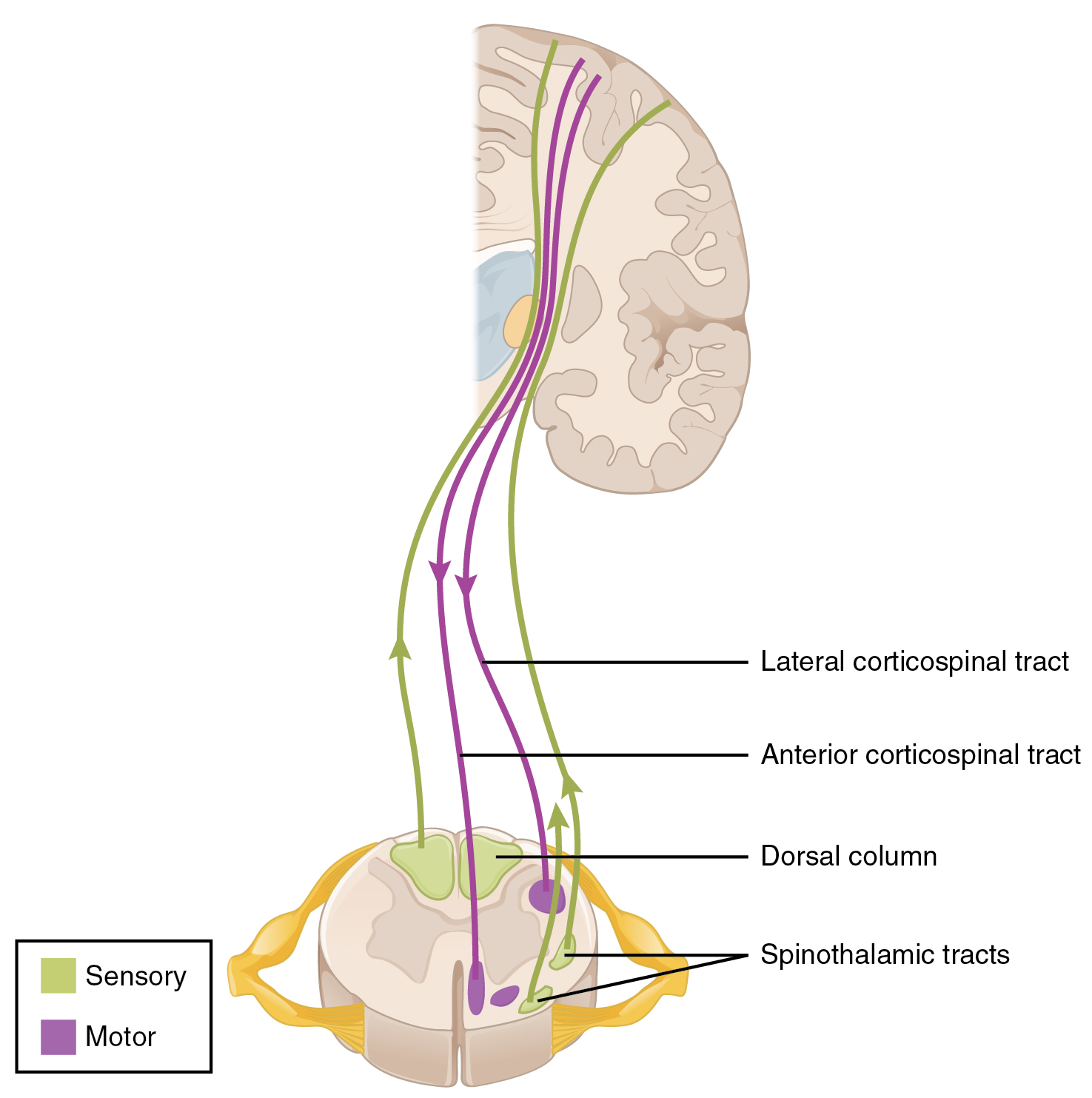
The dorsal column system (sometimes referred to as the dorsal column–medial lemniscus) and the spinothalamic tract are two major pathways that bring sensory information to the brain (Figure 14.5.1). The sensory pathways in each of these systems are composed of three successive neurons.
The dorsal column system begins with the axon of a dorsal root ganglion neuron entering the dorsal root and joining the dorsal column white matter in the spinal cord. As axons of this pathway enter the dorsal column, they take on a positional arrangement so that axons from lower levels of the body position themselves medially, whereas axons from upper levels of the body position themselves laterally. The dorsal column is separated into two component tracts, the fasciculus gracilis that contains axons from the legs and lower body, and the fasciculus cuneatus that contains axons from the upper body and arms.
The axons in the dorsal column terminate in the nuclei of the medulla, where each synapses with the second neuron in their respective pathway. The nucleus gracilis is the target of fibers in the fasciculus gracilis, whereas the nucleus cuneatus is the target of fibers in the fasciculus cuneatus. The second neuron in the system projects from one of the two nuclei and then decussates, or crosses the midline of the medulla. These axons then continue to ascend the brain stem as a bundle called the medial lemniscus. These axons terminate in the thalamus, where each synapses with the third neuron in their respective pathway. The third neuron in the system projects its axons to the postcentral gyrus of the cerebral cortex, where somatosensory stimuli are initially processed and the conscious perception of the stimulus occurs.
The spinothalamic tract also begins with neurons in a dorsal root ganglion. These neurons extend their axons to the dorsal horn, where they synapse with the second neuron in their respective pathway. The name “spinothalamic” comes from this second neuron, which has its cell body in the spinal cord gray matter and connects to the thalamus. Axons from these second neurons then decussate within the spinal cord and ascend to the brain and enter the thalamus, where each synapses with the third neuron in its respective pathway. The neurons in the thalamus then project their axons to the spinothalamic tract, which synapses in the postcentral gyrus of the cerebral cortex.
These two systems are similar in that they both begin with dorsal root ganglion cells, as with most general sensory information. The dorsal column system is primarily responsible for touch sensations and proprioception, whereas the spinothalamic tract pathway is primarily responsible for pain and temperature sensations. Another similarity is that the second neurons in both of these pathways are contralateral, because they project across the midline to the other side of the brain or spinal cord. In the dorsal column system, this decussation takes place in the brain stem; in the spinothalamic pathway, it takes place in the spinal cord at the same spinal cord level at which the information entered. The third neurons in the two pathways are essentially the same. In both, the second neuron synapses in the thalamus, and the thalamic neuron projects to the somatosensory cortex.
The trigeminal pathway carries somatosensory information from the face, head, mouth, and nasal cavity. As with the previously discussed nerve tracts, the sensory pathways of the trigeminal pathway each involve three successive neurons. First, axons from the trigeminal ganglion enter the brain stem at the level of the pons. These axons project to one of three locations. The spinal trigeminal nucleus of the medulla receives information similar to that carried by spinothalamic tract, such as pain and temperature sensations. Other axons go to either the chief sensory nucleus in the pons or the mesencephalic nuclei in the midbrain. These nuclei receive information like that carried by the dorsal column system, such as touch, pressure, vibration, and proprioception. Axons from the second neuron decussate and ascend to the thalamus along the trigeminothalamic tract. In the thalamus, each axon synapses with the third neuron in its respective pathway. Axons from the third neuron then project from the thalamus to the primary somatosensory cortex of the cerebrum.
The diencephalon is beneath the cerebrum and includes the thalamus and hypothalamus. In the somatic nervous system, the thalamus is an important relay for communication between the cerebrum and the rest of the nervous system. The hypothalamus has both somatic and autonomic functions. In addition, the hypothalamus communicates with the limbic system, which controls emotions and memory functions.
Sensory input to the thalamus comes from most of the special senses and ascending somatosensory tracts. Each sensory system is relayed through a particular nucleus in the thalamus. The thalamus is a required transfer point for most sensory tracts that reach the cerebral cortex, where conscious sensory perception begins. The one exception to this rule is the olfactory system. The olfactory tract axons from the olfactory bulb project directly to the cerebral cortex, along with the limbic system and hypothalamus.
The thalamus is a collection of several nuclei that can be categorized into three anatomical groups. White matter running through the thalamus defines the three major regions of the thalamus, which are an anterior nucleus, a medial nucleus, and a lateral group of nuclei. The anterior nucleus serves as a relay between the hypothalamus and the emotion and memory-producing limbic system. The medial nuclei serve as a relay for information from the limbic system and basal ganglia to the cerebral cortex. This allows memory creation during learning, but also determines alertness. The special and somatic senses connect to the lateral nuclei, where their information is relayed to the appropriate sensory cortex of the cerebrum.
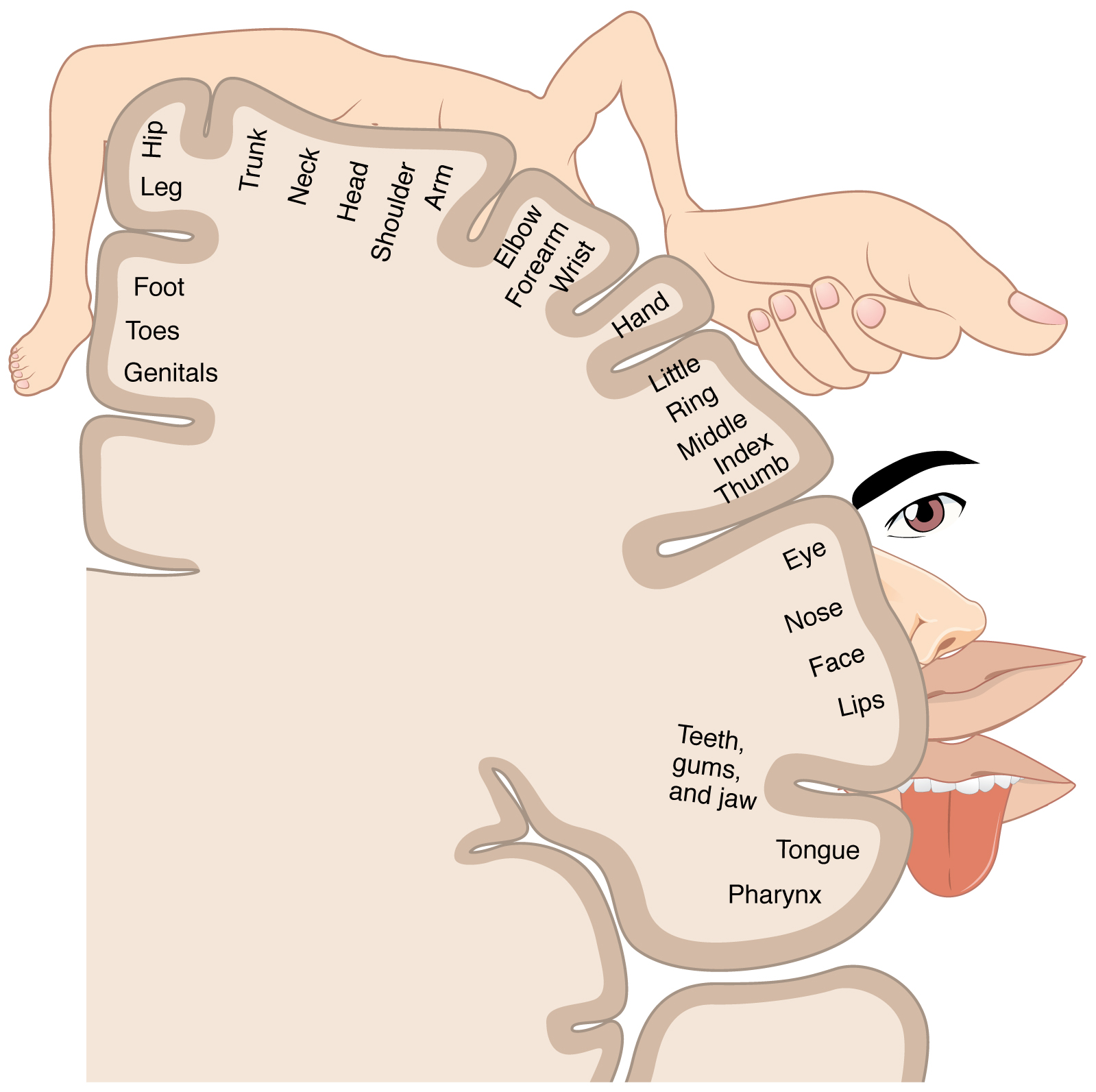
As described earlier, many of the sensory axons are positioned in the same way as their corresponding receptor cells in the body. This allows identification of the position of a stimulus on the basis of which receptor cells are sending information. The cerebral cortex also maintains this sensory topography in the particular areas of the cortex that correspond to the position of the receptor cells. The somatosensory cortex provides an example in which, in essence, the locations of the somatosensory receptors in the body are mapped onto the somatosensory cortex. This mapping is often depicted using a sensory homunculus (Figure 14.5.2).
The term homunculus comes from the Latin word for “little man” and refers to a map of the human body that is laid across a portion of the cerebral cortex. In the somatosensory cortex, the external genitals, feet, and lower legs are represented on the medial face of the gyrus within the longitudinal fissure. As the gyrus curves out of the fissure and along the surface of the parietal lobe, the body map continues through the thighs, hips, trunk, shoulders, arms, and hands. The head and face are just lateral to the fingers as the gyrus approaches the lateral sulcus. The representation of the body in this topographical map is medial to lateral from the lower to upper body. It is a continuation of the topographical arrangement seen in the dorsal column system, where axons from the lower body are carried in the fasciculus gracilis, whereas axons from the upper body are carried in the fasciculus cuneatus. As the dorsal column system continues into the medial lemniscus, these relationships are maintained. Also, the head and neck axons running from the trigeminal nuclei to the thalamus run adjacent to the upper body fibers. The connections through the thalamus maintain topography such that the anatomic information is preserved. Note that this correspondence does not result in a perfectly miniature scale version of the body, but rather exaggerates the more sensitive areas of the body, such as the fingers and lower face. Less sensitive areas of the body, such as the shoulders and back, are mapped to smaller areas on the cortex.
The cortex has been described as having specific regions that are responsible for processing specific information; there is the visual cortex, somatosensory cortex, gustatory cortex, etc. However, our experience of these senses is not divided. Instead, we experience what can be referred to as a seamless percept. Our perceptions of the various sensory modalities—though distinct in their content—are integrated by the brain so that we experience the world as a continuous whole.
In the cerebral cortex, sensory processing begins at the primary sensory cortex, then proceeds to an association area, and finally, into a multimodal integration area. For example, somatosensory information inputs directly into the primary somatosensory cortex in the post-central gyrus of the parietal lobe where general awareness of sensation (location and type of sensation) begins. In the somatosensory association cortex details are integrated into a whole. In the highest level of association cortex details are integrated from entirely different modalities to form complete representations as we experience them.
The defining characteristic of the somatic nervous system is that it controls skeletal muscles. Somatic senses inform the nervous system about the external environment, but the response to that is through voluntary muscle movement. The term “voluntary” suggests that there is a conscious decision to make a movement. However, some aspects of the somatic system use voluntary muscles without conscious control. One example is the ability of our breathing to switch to unconscious control while we are focused on another task. However, the muscles that are responsible for the basic process of breathing are also utilized for speech, which is entirely voluntary.
Let’s start with sensory stimuli that have been registered through receptor cells and the information relayed to the CNS along ascending pathways. In the cerebral cortex, the initial processing of sensory perception progresses to associative processing and then integration in multimodal areas of cortex. These levels of processing can lead to the incorporation of sensory perceptions into memory, but more importantly, they lead to a response. The completion of cortical processing through the primary, associative, and integrative sensory areas initiates a similar progression of motor processing, usually in different cortical areas.
Whereas the sensory cortical areas are located in the occipital, temporal, and parietal lobes, motor functions are largely controlled by the frontal lobe. The most anterior regions of the frontal lobe—the prefrontal areas—are important for executive functions, which are those cognitive functions that lead to goal-directed behaviors. These higher cognitive processes include working memory, which has been called a “mental scratch pad,” that can help organize and represent information that is not in the immediate environment. The prefrontal lobe is responsible for aspects of attention, such as inhibiting distracting thoughts and actions so that a person can focus on a goal and direct behavior toward achieving that goal.
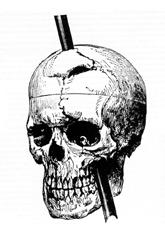
The functions of the prefrontal cortex are integral to the personality of an individual, because it is largely responsible for what a person intends to do and how they accomplish those plans. A famous case of damage to the prefrontal cortex is that of Phineas Gage, dating back to 1848. He was a railroad worker who had a metal spike impale his prefrontal cortex (Figure 14.5.3). He survived the accident, but according to second-hand accounts, his personality changed drastically. Friends described him as no longer acting like himself. Whereas he was a hardworking, amiable man before the accident, he turned into an irritable, temperamental, and lazy man after the accident. Many of the accounts of his change may have been inflated in the retelling, and some behavior was likely attributable to alcohol used as a pain medication. However, the accounts suggest that some aspects of his personality did change. Also, there is new evidence that though his life changed dramatically, he was able to become a functioning stagecoach driver, suggesting that the brain has the ability to recover even from major trauma such as this.

In generating motor responses, the executive functions of the prefrontal cortex will need to initiate actual movements. One way to define the prefrontal area is any region of the frontal lobe that does not elicit movement when electrically stimulated. These are primarily in the anterior part of the frontal lobe. The regions of the frontal lobe that remain are the regions of the cortex that produce movement. The prefrontal areas project into the secondary motor cortices, which include the premotor cortex and the supplemental motor area.
Two important regions that assist in planning and coordinating movements are located adjacent to the primary motor cortex. The premotor cortex is more lateral, whereas the supplemental motor area is more medial and superior. The premotor area aids in controlling movements of the core muscles to maintain posture during movement, whereas the supplemental motor area is hypothesized to be responsible for planning and coordinating movement. The supplemental motor area also manages sequential movements that are based on prior experience (that is, learned movements). Neurons in these areas are most active leading up to the initiation of movement. For example, these areas might prepare the body for the movements necessary to drive a car in anticipation of a traffic light changing.
Adjacent to these two regions are two specialized motor planning centers. The frontal eye fields are responsible for moving the eyes in response to visual stimuli. There are direct connections between the frontal eye fields and the superior colliculus. Also, anterior to the premotor cortex and primary motor cortex is Broca’s area. This area is responsible for controlling movements of the structures of speech production. The area is named after a French surgeon and anatomist who studied patients who could not produce speech. They did not have impairments to understanding speech, only to producing speech sounds, suggesting a damaged or underdeveloped Broca’s area.
The primary motor cortex is located in the precentral gyrus of the frontal lobe. A neurosurgeon, Walter Penfield, described much of the basic understanding of the primary motor cortex by electrically stimulating the surface of the cerebrum. Penfield would probe the surface of the cortex while the patient was only under local anesthesia so that he could observe responses to the stimulation. This led to the belief that the precentral gyrus directly stimulated muscle movement. We now know that the primary motor cortex receives input from several areas that aid in planning movement, and its principle output stimulates spinal cord neurons to stimulate skeletal muscle contraction.
The primary motor cortex is arranged in a similar fashion to the primary somatosensory cortex, in that it has a topographical map of the body, creating a motor homunculus (see Chapter 14.2 Figure 14.2.5). The neurons responsible for musculature in the feet and lower legs are in the medial wall of the precentral gyrus, with the thighs, trunk, and shoulder at the crest of the longitudinal fissure. The hand and face are in the lateral face of the gyrus. Also, the relative space allotted for the different regions is exaggerated in muscles that have greater enervation. The greatest amount of cortical space is given to muscles that perform fine, agile movements, such as the muscles of the fingers and the lower face that are parts of small motor units. The “power muscles” that perform coarser movements, such as the buttock and back muscles, occupy much less space on the motor cortex.
The motor output from the cortex descends into the brain stem and to the spinal cord to control the musculature through motor neurons. Neurons located in the primary motor cortex, named Betz cells, are large cortical neurons that synapse with lower motor neurons in the spinal cord or the brain stem. The two descending pathways travelled by the axons of Betz cells are the corticospinal tract and the corticobulbar tract. Both tracts are named for their origin in the cortex and their targets—either the spinal cord or the brain stem (the term “bulbar” refers to the brain stem as the bulb, or enlargement, at the top of the spinal cord).
These two descending pathways are responsible for the conscious or voluntary movements of skeletal muscles. Any motor command from the primary motor cortex is sent down the axons of the Betz cells to activate upper motor neurons in either the cranial motor nuclei or in the ventral horn of the spinal cord. The axons of the corticobulbar tract are ipsilateral, meaning they project from the cortex to the motor nucleus on the same side of the nervous system. Conversely, the axons of the corticospinal tract are largely contralateral, meaning that they cross the midline of the brain stem or spinal cord and synapse on the opposite side of the body. Therefore, the right motor cortex of the cerebrum controls muscles on the left side of the body, and vice versa.
The corticospinal tract descends from the cortex through the deep white matter of the cerebrum. It then passes between the caudate nucleus and putamen of the basal nuclei as a bundle called the internal capsule. The tract then passes through the midbrain as the cerebral peduncles, after which it burrows through the pons. Upon entering the medulla, the tracts make up the large white matter tract referred to as the pyramids (Figure 14.5.4). The defining landmark of the medullary-spinal border is the pyramidal decussation, which is where most of the fibers in the corticospinal tract cross over to the opposite side of the brain. At this point, the tract separates into two parts, which have control over different domains of the musculature.
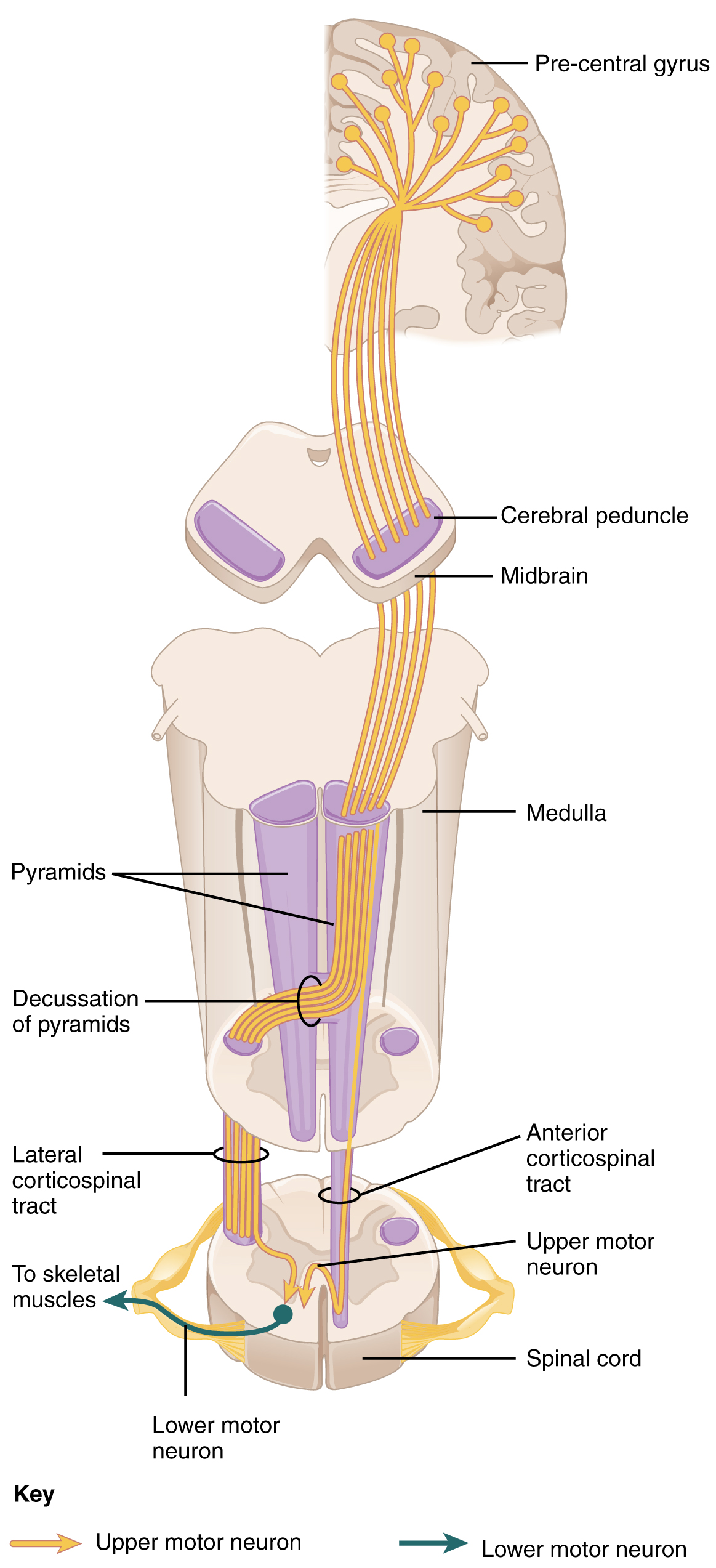
The lateral corticospinal tract is composed of the fibers that cross the midline at the pyramidal decussation (see Figure 14.5.4). The axons cross over from the anterior position of the pyramids in the medulla to the lateral column of the spinal cord. These axons are responsible for controlling appendicular muscles.
This influence over the appendicular muscles means that the lateral corticospinal tract is responsible for moving the muscles of the arms and legs. The ventral horn in both the lower cervical spinal cord and the lumbar spinal cord both have wider ventral horns, representing the greater number of muscles controlled by these motor neurons. The cervical enlargement is particularly large because there is greater control over the fine musculature of the upper limbs, particularly of the fingers. The lumbar enlargement is not as significant in appearance because there is less fine motor control of the lower limbs.
The anterior corticospinal tract is responsible for controlling the muscles of the body trunk (see Figure 14.5.4). These axons do not decussate in the medulla. Instead, they remain in an anterior position as they descend the brain stem and enter the spinal cord. These axons then travel to the spinal cord level at which they synapse with a lower motor neuron. Upon reaching the appropriate level, the axons decussate, entering the ventral horn on the opposite side of the spinal cord from which they entered. In the ventral horn, these axons synapse with their corresponding lower motor neurons. The lower motor neurons are located in the medial regions of the ventral horn, because they control the axial muscles of the trunk.
Because movements of the body trunk involve both sides of the body, the anterior corticospinal tract is not entirely contralateral. Some collateral branches of the tract will project into the ipsilateral ventral horn to control synergistic muscles on that side of the body, or to inhibit antagonistic muscles through interneurons within the ventral horn. Through the influence of both sides of the body, the anterior corticospinal tract can coordinate postural muscles in broad movements of the body. These coordinating axons in the anterior corticospinal tract are often considered bilateral, as they are both ipsilateral and contralateral.
Watch this video to learn more about the descending motor pathway for the somatic nervous system. The autonomic connections are mentioned, which are covered in another chapter. From this brief video, only some of the descending motor pathway of the somatic nervous system is described. Which division of the pathway is described and which division is left out?
Other descending connections between the brain and the spinal cord are called the extrapyramidal system. The name comes from the fact that this system is outside the corticospinal pathway, which includes the pyramids in the medulla. A few pathways originating from the brain stem contribute to this system.
The tectospinal tract projects from the midbrain to the spinal cord and is important for postural movements that are driven by the superior colliculus. The name of the tract comes from an alternate name for the superior colliculus, which is the tectum. The reticulospinal tract connects the reticular system, a diffuse region of gray matter in the brain stem, with the spinal cord. This tract influences trunk and proximal limb muscles related to posture and locomotion. The reticulospinal tract also contributes to muscle tone and influences autonomic functions. The vestibulospinal tract connects the brain stem nuclei of the vestibular system with the spinal cord. This allows posture, movement, and balance to be modulated on the basis of equilibrium information provided by the vestibular system.
The pathways of the extrapyramidal system are influenced by subcortical structures. For example, connections between the secondary motor cortices and the extrapyramidal system modulate spine and cranium movements. The basal nuclei, which are important for regulating movement initiated by the CNS, influence the extrapyramidal system as well as its thalamic feedback to the motor cortex.
The conscious movement of our muscles is more complicated than simply sending a single command from the precentral gyrus down to the proper motor neurons. During the movement of any body part, our muscles relay information back to the brain, and the brain is constantly sending “revised” instructions back to the muscles. The cerebellum is important in contributing to the motor system because it compares cerebral motor commands with proprioceptive feedback. The corticospinal fibers that project to the ventral horn of the spinal cord have branches that also synapse in the pons, which project to the cerebellum. Also, the proprioceptive sensations of the dorsal column system have a collateral projection to the medulla that projects to the cerebellum. These two streams of information are compared in the cerebellar cortex. Conflicts between the motor commands sent by the cerebrum and body position information provided by the proprioceptors cause the cerebellum to stimulate the red nucleus of the midbrain. The red nucleus then sends corrective commands to the spinal cord along the rubrospinal tract. The name of this tract comes from the word for red that is seen in the English word “ruby.”
A good example of how the cerebellum corrects cerebral motor commands can be illustrated by walking in water. An original motor command from the cerebrum to walk will result in a highly coordinated set of learned movements. However, in water, the body cannot actually perform a typical walking movement as instructed. The cerebellum can alter the motor command, stimulating the leg muscles to take larger steps to overcome the water resistance. The cerebellum can make the necessary changes through the rubrospinal tract. Modulating the basic command to walk also relies on spinal reflexes, but the cerebellum is responsible for calculating the appropriate response. When the cerebellum does not work properly, coordination and balance are severely affected. The most dramatic example of this is during the overconsumption of alcohol. Alcohol inhibits the ability of the cerebellum to interpret proprioceptive feedback, making it more difficult to coordinate body movements, such as walking a straight line, or guide the movement of the hand to touch the tip of the nose.
Visit this site to read about an elderly woman who starts to lose the ability to control fine movements, such as speech and the movement of limbs. Many of the usual causes were ruled out. It was not a stroke, Parkinson’s disease, diabetes, or thyroid dysfunction. The next most obvious cause was medication, so her pharmacist had to be consulted. The side effect of a drug meant to help her sleep had resulted in changes in motor control. What regions of the nervous system are likely to be the focus of haloperidol side effects?
Connections between the body and the CNS occur through the spinal cord. The cranial nerves connect the head and neck directly to the brain, but the spinal cord receives sensory input and sends motor commands out to the body through the spinal nerves. Whereas the brain develops into a complex series of nuclei and fiber tracts, the spinal cord remains relatively simple in its configuration (Figure 14.5.5). From the initial neural tube early in embryonic development, the spinal cord retains a tube-like structure with gray matter surrounding the small central canal and white matter on the surface in three columns. The dorsal, or posterior, horns of the gray matter are mainly devoted to sensory functions whereas the ventral, or anterior, and lateral horns are associated with motor functions. In the white matter, the dorsal column relays sensory information to the brain, and the anterior column is almost exclusively relaying motor commands to the ventral horn motor neurons. The lateral column, however, conveys both sensory and motor information between the spinal cord and brain.
The general senses are distributed throughout the body, relying on nervous tissue incorporated into various organs. Somatic senses are incorporated mostly into the skin, muscles, or tendons, whereas the visceral senses come from nervous tissue incorporated into the majority of organs such as the heart or stomach. The somatic senses are those that usually make up the conscious perception of the how the body interacts with the environment. The visceral senses are most often below the limit of conscious perception because they are involved in homeostatic regulation through the autonomic nervous system.
The sensory exam tests the somatic senses, meaning those that are consciously perceived. Testing of the senses begins with examining the regions known as dermatomes that connect to the cortical region where somatosensation is perceived in the postcentral gyrus. To test the sensory fields, a simple stimulus of the light touch of the soft end of a cotton-tipped applicator is applied at various locations on the skin. The spinal nerves, which contain sensory fibers with dendritic endings in the skin, connect with the skin in a topographically organized manner, illustrated as dermatomes (Figure 14.5.6). For example, the fibers of eighth cervical nerve innervate the medial surface of the forearm and extend out to the fingers. In addition to testing perception at different positions on the skin, it is necessary to test sensory perception within the dermatome from distal to proximal locations in the appendages, or lateral to medial locations in the trunk. In testing the eighth cervical nerve, the patient would be asked if the touch of the cotton to the fingers or the medial forearm was perceptible, and whether there were any differences in the sensations.
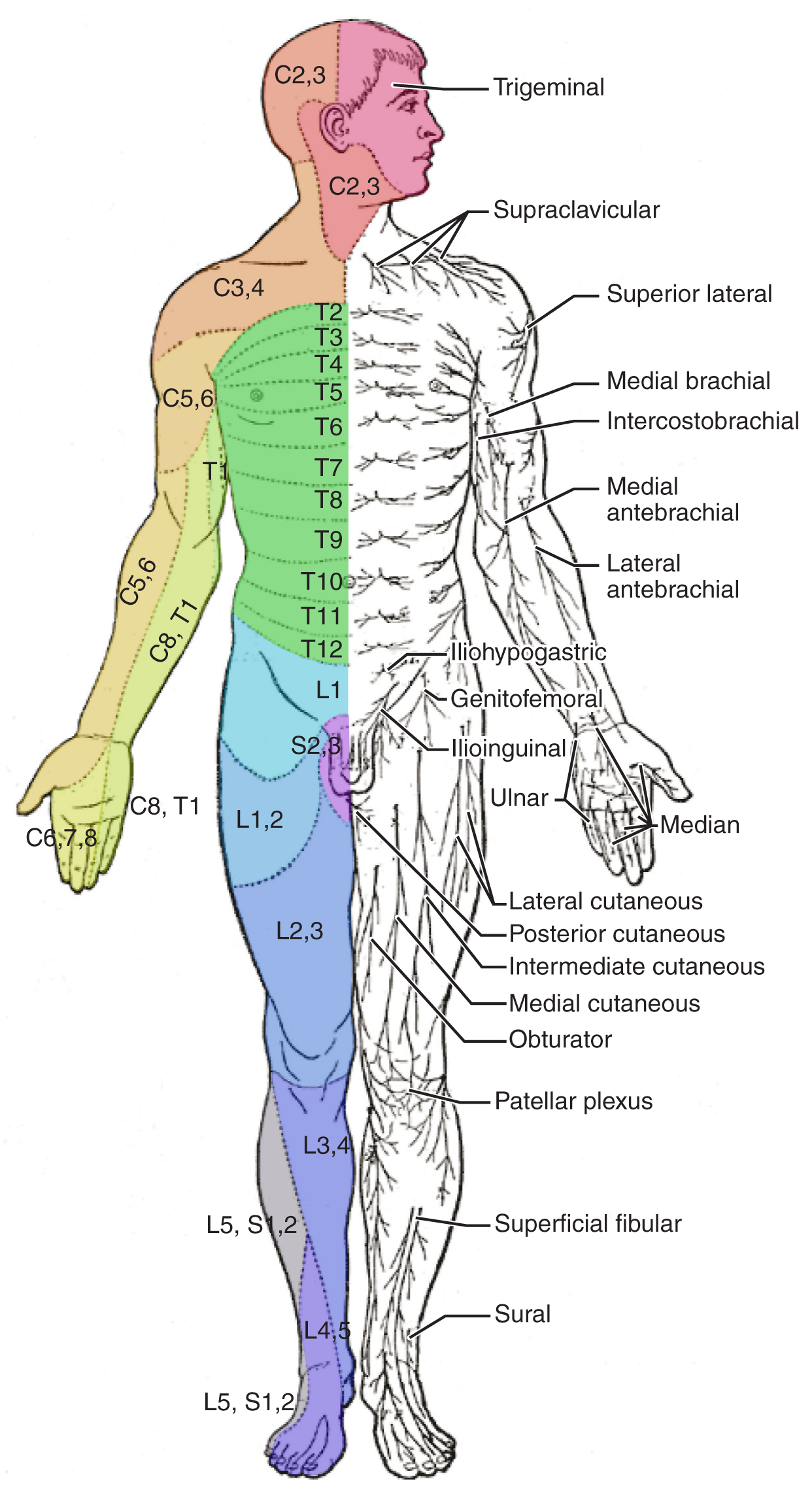
Other modalities of somatosensation can be tested using a few simple tools. The perception of pain can be tested using the broken end of the cotton-tipped applicator. The perception of vibratory stimuli can be testing using an oscillating tuning fork placed against prominent bone features such as the distal head of the ulna on the medial aspect of the elbow. When the tuning fork is still, the metal against the skin can be perceived as a cold stimulus. Using the cotton tip of the applicator, or even just a fingertip, the perception of tactile movement can be assessed as the stimulus is drawn across the skin for approximately 2–3 cm. The patient would be asked in what direction the stimulus is moving. All of these tests are repeated in distal and proximal locations and for different dermatomes to assess the spatial specificity of perception. The sense of position and motion, proprioception, is tested by moving the fingers or toes and asking the patient if they sense the movement. If the distal locations are not perceived, the test is repeated at increasingly proximal joints.
The various stimuli used to test sensory input assess the function of the major ascending tracts of the spinal cord. The dorsal column pathway conveys fine touch, vibration, and proprioceptive information, whereas the spinothalamic pathway primarily conveys pain and temperature. Testing these stimuli provides information about whether these two major ascending pathways are functioning properly. Within the spinal cord, the two systems are segregated. The dorsal column information ascends ipsilateral to the source of the stimulus and decussates in the medulla, whereas the spinothalamic pathway decussates at the level of entry and ascends contralaterally. The differing sensory stimuli are segregated in the spinal cord so that the various subtests for these stimuli can distinguish which ascending pathway may be damaged in certain situations.
Whereas the basic sensory stimuli are assessed in the subtests directed at each submodality of somatosensation, testing the ability to discriminate sensations is important. Pairing the light touch and pain subtests together makes it possible to compare the two submodalities at the same time, and therefore the two major ascending tracts at the same time. Mistaking painful stimuli for light touch, or vice versa, may point to errors in ascending projections, such as in a hemisection of the spinal cord that might come from a motor vehicle accident.
Another issue of sensory discrimination is not distinguishing between different submodalities, but rather location. The two-point discrimination subtest highlights the density of sensory endings, and therefore receptive fields in the skin. The sensitivity to fine touch, which can give indications of the texture and detailed shape of objects, is highest in the fingertips. To assess the limit of this sensitivity, two-point discrimination is measured by simultaneously touching the skin in two locations, such as could be accomplished with a pair of forceps. Specialized calipers for precisely measuring the distance between points are also available. The patient is asked to indicate whether one or two stimuli are present while keeping their eyes closed. The examiner will switch between using the two points and a single point as the stimulus. Failure to recognize two points may be an indication of a dorsal column pathway deficit.
Similar to two-point discrimination, but assessing laterality of perception, is double simultaneous stimulation. Two stimuli, such as the cotton tips of two applicators, are touched to the same position on both sides of the body. If one side is not perceived, this may indicate damage to the contralateral posterior parietal lobe. Because there is one of each pathway on either side of the spinal cord, they are not likely to interact. If none of the other subtests suggest particular deficits with the pathways, the deficit is likely to be in the cortex where conscious perception is based. The mental status exam contains subtests that assess other functions that are primarily localized to the parietal cortex, such as stereognosis and graphesthesia.
A final subtest of sensory perception that concentrates on the sense of proprioception is known as the Romberg test. The patient is asked to stand straight with feet together. Once the patient has achieved their balance in that position, they are asked to close their eyes. Without visual feedback that the body is in a vertical orientation relative to the surrounding environment, the patient must rely on the proprioceptive stimuli of joint and muscle position, as well as information from the inner ear, to maintain balance. This test can indicate deficits in dorsal column pathway proprioception, as well as problems with proprioceptive projections to the cerebellum through the spinocerebellar tract.
Watch this video to see a quick demonstration of two-point discrimination. Touching a specialized caliper to the surface of the skin will measure the distance between two points that are perceived as distinct stimuli versus a single stimulus. The patient keeps their eyes closed while the examiner switches between using both points of the caliper or just one. The patient then must indicate whether one or two stimuli are in contact with the skin. Why is the distance between the caliper points closer on the fingertips as opposed to the palm of the hand? And what do you think the distance would be on the arm, or the shoulder?
The skeletomotor system is largely based on the simple, two-cell projection from the precentral gyrus of the frontal lobe to the skeletal muscles. The corticospinal tract represents the neurons that send output from the primary motor cortex. These fibers travel through the deep white matter of the cerebrum, then through the midbrain and pons, into the medulla where most of them decussate, and finally through the spinal cord white matter in the lateral (crossed fibers) or anterior (uncrossed fibers) columns. These fibers synapse on motor neurons in the ventral horn. The ventral horn motor neurons then project to skeletal muscle and cause contraction. These two cells are termed the upper motor neuron (UMN) and the lower motor neuron (LMN). Voluntary movements require these two cells to be active.
The motor exam tests the function of these neurons and the muscles they control. First, the muscles are inspected and palpated for signs of structural irregularities. Movement disorders may be the result of changes to the muscle tissue, such as scarring, and these possibilities need to be ruled out before testing function. Along with this inspection, muscle tone is assessed by moving the muscles through a passive range of motion. The arm is moved at the elbow and wrist, and the leg is moved at the knee and ankle. Skeletal muscle should have a resting tension representing a slight contraction of the fibers. The lack of muscle tone, known as hypotonicity or flaccidity, may indicate that the LMN is not conducting action potentials that will keep a basal level of acetylcholine in the neuromuscular junction.
If muscle tone is present, muscle strength is tested by having the patient contract muscles against resistance. The examiner will ask the patient to lift the arm, for example, while the examiner is pushing down on it. This is done for both limbs, including shrugging the shoulders. Lateral differences in strength—being able to push against resistance with the right arm but not the left—would indicate a deficit in one corticospinal tract versus the other. An overall loss of strength, without laterality, could indicate a global problem with the motor system. Diseases that result in UMN lesions include cerebral palsy or MS, or it may be the result of a stroke. A sign of UMN lesion is a negative result in the subtest for pronator drift. The patient is asked to extend both arms in front of the body with the palms facing up. While keeping the eyes closed, if the patient unconsciously allows one or the other arm to slowly relax, toward the pronated position, this could indicate a failure of the motor system to maintain the supinated position.
Reflexes combine the spinal sensory and motor components with a sensory input that directly generates a motor response. The reflexes that are tested in the neurological exam are classified into two groups. A deep tendon reflex is commonly known as a stretch reflex, and is elicited by a strong tap to a tendon, such as in the knee-jerk reflex. A superficial reflex is elicited through gentle stimulation of the skin and causes contraction of the associated muscles.For the arm, the common reflexes to test are of the biceps, brachioradialis, triceps, and flexors for the digits. For the leg, the knee-jerk reflex of the quadriceps is common, as is the ankle reflex for the gastrocnemius and soleus. The tendon at the insertion for each of these muscles is struck with a rubber mallet. The muscle is quickly stretched, resulting in activation of the muscle spindle that sends a signal into the spinal cord through the dorsal root. The fiber synapses directly on the ventral horn motor neuron that activates the muscle, causing contraction. The reflexes are physiologically useful for stability. If a muscle is stretched, it reflexively contracts to return the muscle to compensate for the change in length. In the context of the neurological exam, reflexes indicate that the LMN is functioning properly.The most common superficial reflex in the neurological exam is the plantar reflex that tests for the Babinski sign on the basis of the extension or flexion of the toes at the plantar surface of the foot. The plantar reflex is commonly tested in newborn infants to establish the presence of neuromuscular function. To elicit this reflex, an examiner brushes a stimulus, usually the examiner’s fingertip, along the plantar surface of the infant’s foot. An infant would present a positive Babinski sign, meaning the foot dorsiflexes and the toes extend and splay out. As a person learns to walk, the plantar reflex changes to cause curling of the toes and a moderate plantar flexion. If superficial stimulation of the sole of the foot caused extension of the foot, keeping one’s balance would be harder. The descending input of the corticospinal tract modifies the response of the plantar reflex, meaning that a negative Babinski sign is the expected response in testing the reflex. Other superficial reflexes are not commonly tested, though a series of abdominal reflexes can target function in the lower thoracic spinal segments.
Watch this video to see how to test reflexes in the abdomen. Testing reflexes of the trunk is not commonly performed in the neurological exam, but if findings suggest a problem with the thoracic segments of the spinal cord, a series of superficial reflexes of the abdomen can localize function to those segments. If contraction is not observed when the skin lateral to the umbilicus (belly button) is stimulated, what level of the spinal cord may be damaged?
Many of the tests of motor function can indicate differences that will address whether damage to the motor system is in the upper or lower motor neurons. Signs that suggest a UMN lesion include muscle weakness, strong deep tendon reflexes, decreased control of movement or slowness, pronator drift, a positive Babinski sign, spasticity, and the clasp-knife response. Spasticity is an excess contraction in resistance to stretch. It can result in hyperflexia, which is when joints are overly flexed. The clasp-knife response occurs when the patient initially resists movement, but then releases, and the joint will quickly flex like a pocket knife closing.
A lesion on the LMN would result in paralysis, or at least partial loss of voluntary muscle control, which is known as paresis. The paralysis observed in LMN diseases is referred to as flaccid paralysis, referring to a complete or partial loss of muscle tone, in contrast to the loss of control in UMN lesions in which tone is retained and spasticity is exhibited. Other signs of an LMN lesion are fibrillation, fasciculation, and compromised or lost reflexes resulting from the denervation of the muscle fibers.
Disorders of the…Spinal Cord
In certain situations, such as a motorcycle accident, only half of the spinal cord may be damaged in what is known as a hemisection. Forceful trauma to the trunk may cause ribs or vertebrae to fracture, and debris can crush or section through part of the spinal cord. The full section of a spinal cord would result in paraplegia, or loss of voluntary motor control of the lower body, as well as loss of sensations from that point down. A hemisection, however, will leave spinal cord tracts intact on one side. The resulting condition would be hemiplegia on the side of the trauma—one leg would be paralyzed. The sensory results are more complicated.
The ascending tracts in the spinal cord are segregated between the dorsal column and spinothalamic pathways. This means that the sensory deficits will be based on the particular sensory information each pathway conveys. Sensory discrimination between touch and painful stimuli will illustrate the difference in how these pathways divide these functions.
On the paralyzed leg, a patient will acknowledge painful stimuli, but not fine touch or proprioceptive sensations. On the functional leg, the opposite is true. The reason for this is that the dorsal column pathway ascends ipsilateral to the sensation, so it would be damaged the same way as the lateral corticospinal tract. The spinothalamic pathway decussates immediately upon entering the spinal cord and ascends contralateral to the source; it would therefore bypass the hemisection.
The motor system can indicate the loss of input to the ventral horn in the lumbar enlargement where motor neurons to the leg are found, but motor function in the trunk is less clear. The left and right anterior corticospinal tracts are directly adjacent to each other. The likelihood of trauma to the spinal cord resulting in a hemisection that affects one anterior column, but not the other, is very unlikely. Either the axial musculature will not be affected at all, or there will be bilateral losses in the trunk.
Sensory discrimination can pinpoint the level of damage in the spinal cord. Below the hemisection, pain stimuli will be perceived in the damaged side, but not fine touch. The opposite is true on the other side. The pain fibers on the side with motor function cross the midline in the spinal cord and ascend in the contralateral lateral column as far as the hemisection. The dorsal column will be intact ipsilateral to the source on the intact side and reach the brain for conscious perception. The trauma would be at the level just before sensory discrimination returns to normal, helping to pinpoint the trauma. Whereas imaging technology, like magnetic resonance imaging (MRI) or computed tomography (CT) scanning, could localize the injury as well, nothing more complicated than a cotton-tipped applicator can localize the damage. That may be all that is available on the scene when moving the victim requires crucial decisions be made.
The cerebellum is located in apposition to the dorsal surface of the brain stem, centered on the pons. The name of the pons is derived from its connection to the cerebellum. The word means “bridge” and refers to the thick bundle of myelinated axons that form a bulge on its ventral surface. Those fibers are axons that project from the gray matter of the pons into the contralateral cerebellar cortex. These fibers make up the middle cerebellar peduncle (MCP) and are the major physical connection of the cerebellum to the brain stem (Figure 14.5.7). Two other white matter bundles connect the cerebellum to the other regions of the brain stem. The superior cerebellar peduncle (SCP) is the connection of the cerebellum to the midbrain and forebrain. The inferior cerebellar peduncle (ICP) is the connection to the medulla.
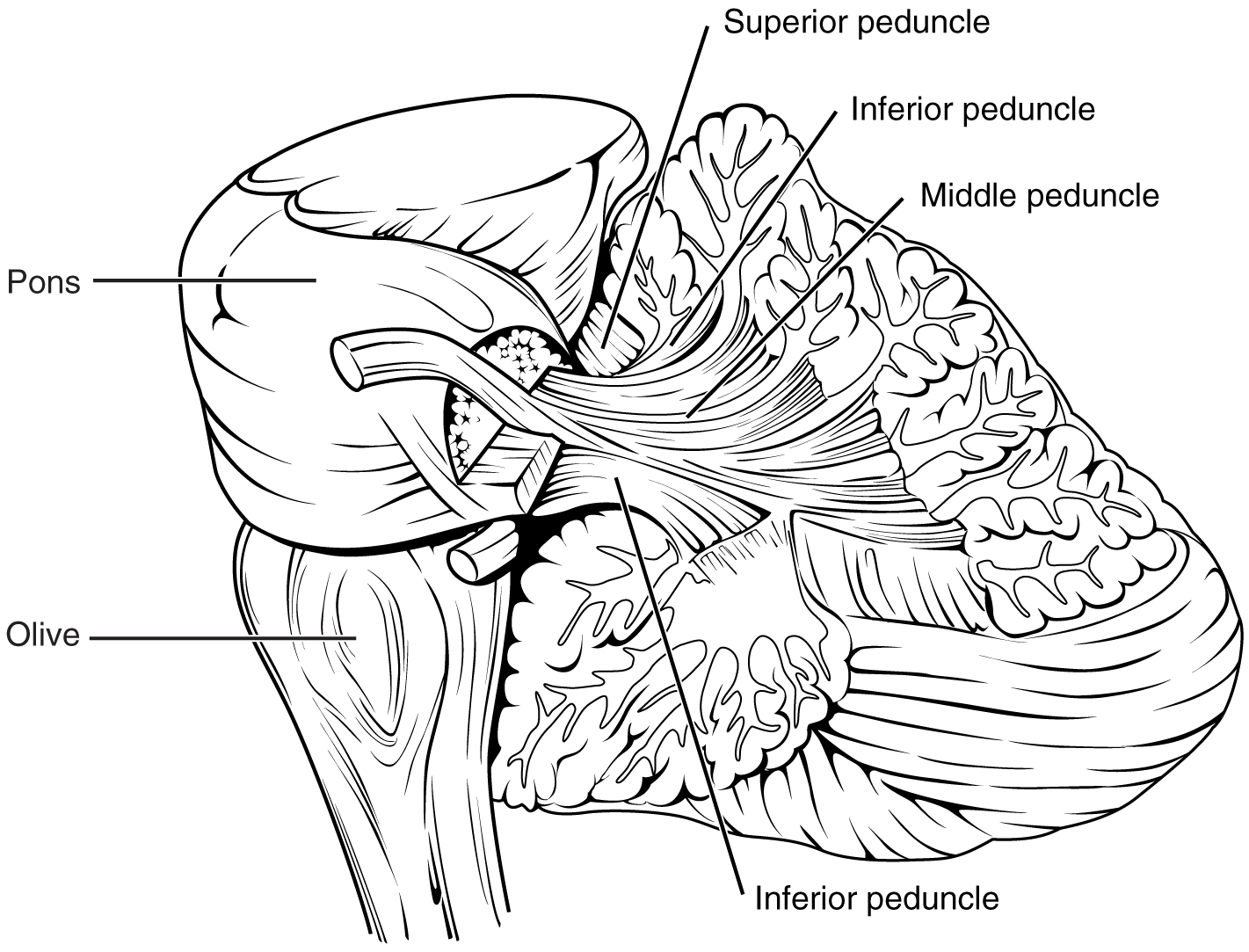
These connections can also be broadly described by their functions. The ICP conveys sensory input to the cerebellum, partially from the spinocerebellar tract, but also through fibers of the inferior olive. The MCP is part of the cortico-ponto-cerebellar pathway that connects the cerebral cortex with the cerebellum and preferentially targets the lateral regions of the cerebellum. It includes a copy of the motor commands sent from the precentral gyrus through the corticospinal tract, arising from collateral branches that synapse in the gray matter of the pons, along with input from other regions such as the visual cortex. The SCP is the major output of the cerebellum, divided between the red nucleus in the midbrain and the thalamus, which will return cerebellar processing to the motor cortex. These connections describe a circuit that compares motor commands and sensory feedback to generate a new output. These comparisons make it possible to coordinate movements. If the cerebral cortex sends a motor command to initiate walking, that command is copied by the pons and sent into the cerebellum through the MCP. Sensory feedback in the form of proprioception from the spinal cord, as well as vestibular sensations from the inner ear, enters through the ICP. If you take a step and begin to slip on the floor because it is wet, the output from the cerebellum—through the SCP—can correct for that and keep you balanced and moving. The red nucleus sends new motor commands to the spinal cord through the rubrospinal tract.
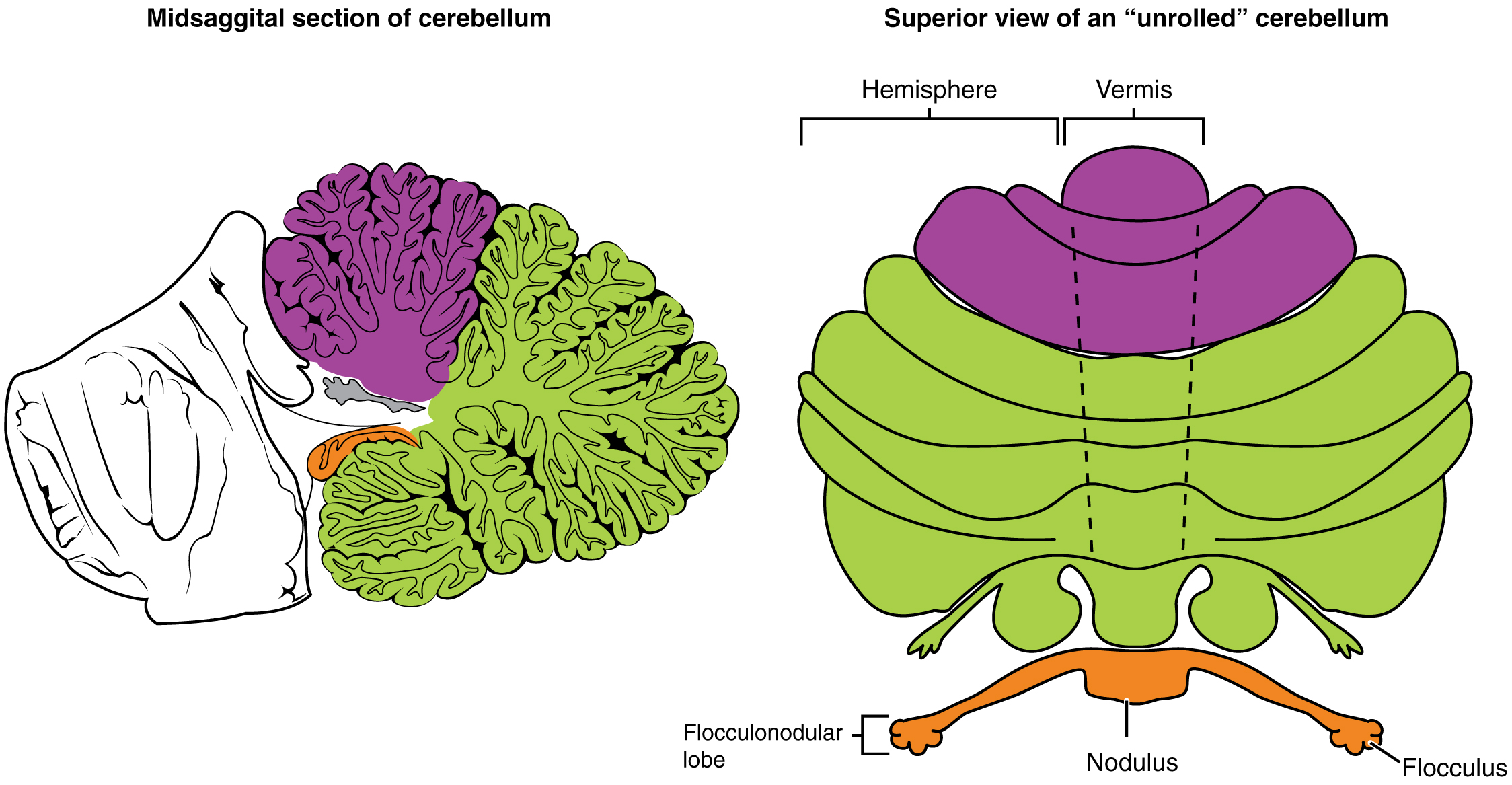
The cerebellum is divided into regions that are based on the particular functions and connections involved. The midline regions of the cerebellum, the vermis and flocculonodular lobe, are involved in comparing visual information, equilibrium, and proprioceptive feedback to maintain balance and coordinate movements such as walking, or gait, through the descending output of the red nucleus (Figure 15.5.8). The lateral hemispheres are primarily concerned with planning motor functions through frontal lobe inputs that are returned through the thalamic projections back to the premotor and motor cortices. Processing in the midline regions targets movements of the axial musculature, whereas the lateral regions target movements of the appendicular musculature. The vermis is referred to as the spinocerebellum because it primarily receives input from the dorsal columns and spinocerebellar pathways. The flocculonodular lobe is referred to as the vestibulocerebellum because of the vestibular projection into that region. Finally, the lateral cerebellum is referred to as the cerebrocerebellum, reflecting the significant input from the cerebral cortex through the cortico-ponto-cerebellar pathway.
Testing for cerebellar function is the basis of the coordination exam. The subtests target appendicular musculature, controlling the limbs, and axial musculature for posture and gait. The assessment of cerebellar function will depend on the normal functioning of other systems addressed in previous sections of the neurological exam. Motor control from the cerebrum, as well as sensory input from somatic, visual, and vestibular senses, are important to cerebellar function.
The subtests that address appendicular musculature, and therefore the lateral regions of the cerebellum, begin with a check for tremor. The patient extends their arms in front of them and holds the position. The examiner watches for the presence of tremors that would not be present if the muscles are relaxed. By pushing down on the arms in this position, the examiner can check for the rebound response, which is when the arms are automatically brought back to the extended position. The extension of the arms is an ongoing motor process, and the tap or push on the arms presents a change in the proprioceptive feedback. The cerebellum compares the cerebral motor command with the proprioceptive feedback and adjusts the descending input to correct. The red nucleus would send an additional signal to the LMN for the arm to increase contraction momentarily to overcome the change and regain the original position.
The check reflex depends on cerebellar input to keep increased contraction from continuing after the removal of resistance. The patient flexes the elbow against resistance from the examiner to extend the elbow. When the examiner releases the arm, the patient should be able to stop the increased contraction and keep the arm from moving. A similar response would be seen if you try to pick up a coffee mug that you believe to be full but turns out to be empty. Without checking the contraction, the mug would be thrown from the overexertion of the muscles expecting to lift a heavier object.
Several subtests of the cerebellum assess the ability to alternate movements, or switch between muscle groups that may be antagonistic to each other. In the finger-to-nose test, the patient touches their finger to the examiner’s finger and then to their nose, and then back to the examiner’s finger, and back to the nose. The examiner moves the target finger to assess a range of movements. A similar test for the lower extremities has the patient touch their toe to a moving target, such as the examiner’s finger. Both of these tests involve flexion and extension around a joint—the elbow or the knee and the shoulder or hip—as well as movements of the wrist and ankle. The patient must switch between the opposing muscles, like the biceps and triceps brachii, to move their finger from the target to their nose. Coordinating these movements involves the motor cortex communicating with the cerebellum through the pons and feedback through the thalamus to plan the movements. Visual cortex information is also part of the processing that occurs in the cerebrocerebellum while it is involved in guiding movements of the finger or toe.
Rapid, alternating movements are tested for the upper and lower extremities. The patient is asked to touch each finger to their thumb, or to pat the palm of one hand on the back of the other, and then flip that hand over and alternate back-and-forth. To test similar function in the lower extremities, the patient touches their heel to their shin near the knee and slides it down toward the ankle, and then back again, repetitively. Rapid, alternating movements are part of speech as well. A patient is asked to repeat the nonsense consonants “lah-kah-pah” to alternate movements of the tongue, lips, and palate. All of these rapid alternations require planning from the cerebrocerebellum to coordinate movement commands that control the coordination.
Gait can either be considered a separate part of the neurological exam or a subtest of the coordination exam that addresses walking and balance. Testing posture and gait addresses functions of the spinocerebellum and the vestibulocerebellum because both are part of these activities. A subtest called station begins with the patient standing in a normal position to check for the placement of the feet and balance. The patient is asked to hop on one foot to assess the ability to maintain balance and posture during movement. Though the station subtest appears to be similar to the Romberg test, the difference is that the patient’s eyes are open during station. The Romberg test has the patient stand still with the eyes closed. Any changes in posture would be the result of proprioceptive deficits, and the patient is able to recover when they open their eyes.
Subtests of walking begin with having the patient walk normally for a distance away from the examiner, and then turn and return to the starting position. The examiner watches for abnormal placement of the feet and the movement of the arms relative to the movement. The patient is then asked to walk with a few different variations. Tandem gait is when the patient places the heel of one foot against the toe of the other foot and walks in a straight line in that manner. Walking only on the heels or only on the toes will test additional aspects of balance.
A movement disorder of the cerebellum is referred to as ataxia. It presents as a loss of coordination in voluntary movements. Ataxia can also refer to sensory deficits that cause balance problems, primarily in proprioception and equilibrium. When the problem is observed in movement, it is ascribed to cerebellar damage. Sensory and vestibular ataxia would likely also present with problems in gait and station.
Ataxia is often the result of exposure to exogenous substances, focal lesions, or a genetic disorder. Focal lesions include strokes affecting the cerebellar arteries, tumors that may impinge on the cerebellum, trauma to the back of the head and neck, or MS. Alcohol intoxication or drugs such as ketamine cause ataxia, but it is often reversible. Mercury in fish can cause ataxia as well. Hereditary conditions can lead to degeneration of the cerebellum or spinal cord, as well as malformation of the brain, or the abnormal accumulation of copper seen in Wilson’s disease.
Watch this short video to see a test for station. Station refers to the position a person adopts when they are standing still. The examiner would look for issues with balance, which coordinates proprioceptive, vestibular, and visual information in the cerebellum. To test the ability of a subject to maintain balance, asking them to stand or hop on one foot can be more demanding. The examiner may also push the subject to see if they can maintain balance. An abnormal finding in the test of station is if the feet are placed far apart. Why would a wide stance suggest problems with cerebellar function?
The neurological exam has been described as a clinical tool throughout this chapter. It is also useful in other ways. A variation of the coordination exam is the Field Sobriety Test (FST) used to assess whether drivers are under the influence of alcohol. The cerebellum is crucial for coordinated movements such as keeping balance while walking, or moving appendicular musculature on the basis of proprioceptive feedback. The cerebellum is also very sensitive to ethanol, the particular type of alcohol found in beer, wine, and liquor.
Walking in a straight line involves comparing the motor command from the primary motor cortex to the proprioceptive and vestibular sensory feedback, as well as following the visual guide of the white line on the side of the road. When the cerebellum is compromised by alcohol, the cerebellum cannot coordinate these movements effectively, and maintaining balance becomes difficult.
Another common aspect of the FST is to have the driver extend their arms out wide and touch their fingertip to their nose, usually with their eyes closed. The point of this is to remove the visual feedback for the movement and force the driver to rely just on proprioceptive information about the movement and position of their fingertip relative to their nose. With eyes open, the corrections to the movement of the arm might be so small as to be hard to see, but proprioceptive feedback is not as immediate and broader movements of the arm will probably be needed, particularly if the cerebellum is affected by alcohol.
Reciting the alphabet backwards is not always a component of the FST, but its relationship to neurological function is interesting. There is a cognitive aspect to remembering how the alphabet goes and how to recite it backwards. That is actually a variation of the mental status subtest of repeating the months backwards. However, the cerebellum is important because speech production is a coordinated activity. The speech rapid alternating movement subtest is specifically using the consonant changes of “lah-kah-pah” to assess coordinated movements of the lips, tongue, pharynx, and palate. But the entire alphabet, especially in the nonrehearsed backwards order, pushes this type of coordinated movement quite far. It is related to the reason that speech becomes slurred when a person is intoxicated.
Sensory input to the brain enters through pathways that travel through either the spinal cord (for somatosensory input from the body) or the brain stem (for everything else, except the visual and olfactory systems) to reach the diencephalon. In the diencephalon, sensory pathways reach the thalamus. This is necessary for all sensory systems to reach the cerebral cortex, except for the olfactory system that is directly connected to the frontal and temporal lobes.
The two major tracts in the spinal cord, originating from sensory neurons in the dorsal root ganglia, are the dorsal column system and the spinothalamic tract. The major differences between the two are in the type of information that is relayed to the brain and where the tracts decussate. The dorsal column system primarily carries information about touch and proprioception and crosses the midline in the medulla. The spinothalamic tract is primarily responsible for pain and temperature sensation and crosses the midline in the spinal cord at the level at which it enters. The trigeminal nerve adds similar sensation information from the head to these pathways.
The motor components of the somatic nervous system begin with the frontal lobe of the brain, where the prefrontal cortex is responsible for higher functions such as working memory. The integrative and associate functions of the prefrontal lobe feed into the secondary motor areas, which help plan movements. The premotor cortex and supplemental motor area then feed into the primary motor cortex that initiates movements. Large Betz cells project through the corticobulbar and corticospinal tracts to synapse on lower motor neurons in the brain stem and ventral horn of the spinal cord, respectively. These connections are responsible for generating movements of skeletal muscles.
The extrapyramidal system includes projections from the brain stem and higher centers that influence movement, mostly to maintain balance and posture, as well as to maintain muscle tone. The superior colliculus and red nucleus in the midbrain, the vestibular nuclei in the medulla, and the reticular formation throughout the brain stem each have tracts projecting to the spinal cord in this system. Descending input from the secondary motor cortices, basal nuclei, and cerebellum connect to the origins of these tracts in the brain stem.
All of these motor pathways project to the spinal cord to synapse with motor neurons in the ventral horn of the spinal cord. These lower motor neurons are the cells that connect to skeletal muscle and cause contractions. These neurons project through the spinal nerves to connect to the muscles at neuromuscular junctions. One motor neuron connects to multiple muscle fibers within a target muscle. The number of fibers that are innervated by a single motor neuron varies on the basis of the precision necessary for that muscle and the amount of force necessary for that motor unit. The quadriceps, for example, have many fibers controlled by single motor neurons for powerful contractions that do not need to be precise. The extraocular muscles have only a small number of fibers controlled by each motor neuron because moving the eyes does not require much force, but needs to be very precise.
Reflexes are the simplest circuits within the somatic nervous system. A withdrawal reflex from a painful stimulus only requires the sensory fiber that enters the spinal cord and the motor neuron that projects to a muscle. Antagonist and postural muscles can be coordinated with the withdrawal, making the connections more complex. The simple, single neuronal connection is the basis of somatic reflexes. The corneal reflex is contraction of the orbicularis oculi muscle to blink the eyelid when something touches the surface of the eye. Stretch reflexes maintain a constant length of muscles by causing a contraction of a muscle to compensate for a stretch that can be sensed by a specialized receptor called a muscle spindle.
Watch this video to learn more about the descending motor pathway for the somatic nervous system. The autonomic connections are mentioned, which are covered in another chapter. From this brief video, only some of the descending motor pathway of the somatic nervous system is described. Which division of the pathway is described and which division is left out?
The video only describes the lateral division of the corticospinal tract. The anterior division is omitted.
Visit this site to read about an elderly woman who starts to lose the ability to control fine movements, such as speech and the movement of limbs. Many of the usual causes were ruled out. It was not a stroke, Parkinson’s disease, diabetes, or thyroid dysfunction. The next most obvious cause was medication, so her pharmacist had to be consulted. The side effect of a drug meant to help her sleep had resulted in changes in motor control. What regions of the nervous system are likely to be the focus of haloperidol side effects?
The movement disorders were similar to those seen in movement disorders of the extrapyramidal system, which would mean the basal nuclei are the most likely source of haloperidol side effects. In fact, haloperidol affects dopamine activity, which is a prominent part of the chemistry of the basal nuclei.
Watch this video to learn more about the reflex arc of the corneal reflex. When the right cornea senses a tactile stimulus, what happens to the left eye? Explain your answer.
The left eye also blinks. The sensory input from one eye activates the motor response of both eyes so that they both blink.
Watch this video to learn more about newborn reflexes. Newborns have a set of reflexes that are expected to have been crucial to survival before the modern age. These reflexes disappear as the baby grows, as some of them may be unnecessary as they age. The video demonstrates a reflex called the Babinski reflex, in which the foot flexes dorsally and the toes splay out when the sole of the foot is lightly scratched. This is normal for newborns, but it is a sign of reduced myelination of the spinal tract in adults. Why would this reflex be a problem for an adult?
While walking, the sole of the foot may be scraped or scratched by many things. If the foot still reacted as in the Babinski reflex, an adult might lose their balance while walking.
ascending pathway fiber structure that relays sensory information from the periphery through the spinal cord and brain stem to other structures of the brain association area region of cortex connected to a primary sensory cortical area that further processes the information to generate more complex sensory perceptions chief sensory nucleus component of the trigeminal nuclei that is found in the pons decussate to cross the midline, as in fibers that project from one side of the body to the other dorsal column system ascending tract of the spinal cord associated with fine touch and proprioceptive sensations fasciculus cuneatus lateral division of the dorsal column system composed of fibers from sensory neurons in the upper body fasciculus gracilis medial division of the dorsal column system composed of fibers from sensory neurons in the lower body medial lemniscus fiber tract of the dorsal column system that extends from the nuclei gracilis and cuneatus to the thalamus, and decussates mesencephalic nucleus component of the trigeminal nuclei that is found in the midbrain multimodal integration area region of the cerebral cortex in which information from more than one sensory modality is processed to arrive at higher level cortical functions such as memory, learning, or cognition nucleus cuneatus medullary nucleus at which first-order neurons of the dorsal column system synapse specifically from the upper body and arms nucleus gracilis medullary nucleus at which first-order neurons of the dorsal column system synapse specifically from the lower body and legs primary sensory cortex region of the cerebral cortex that initially receives sensory input from an ascending pathway from the thalamus and begins the processing that will result in conscious perception of that modality sensory homunculus topographic representation of the body within the somatosensory cortex demonstrating the correspondence between neurons processing stimuli and sensitivity spinal trigeminal nucleus component of the trigeminal nuclei that is found in the medulla spinothalamic tract ascending tract of the spinal cord associated with pain and temperature sensations
anterior corticospinal tract division of the corticospinal pathway that travels through the ventral (anterior) column of the spinal cord and controls axial musculature through the medial motor neurons in the ventral (anterior) horn Betz cells output cells of the primary motor cortex that cause musculature to move through synapses on cranial and spinal motor neurons Broca’s area region of the frontal lobe associated with the motor commands necessary for speech production cerebral peduncles segments of the descending motor pathway that make up the white matter of the ventral midbrain cervical enlargement region of the ventral (anterior) horn of the spinal cord that has a larger population of motor neurons for the greater number of and finer control of muscles of the upper limb corneal reflex protective response to stimulation of the cornea causing contraction of the orbicularis oculi muscle resulting in blinking of the eye corticobulbar tract connection between the cortex and the brain stem responsible for generating movement corticospinal tract connection between the cortex and the spinal cord responsible for generating movement executive functions cognitive processes of the prefrontal cortex that lead to directing goal-directed behavior, which is a precursor to executing motor commands extrapyramidal system pathways between the brain and spinal cord that are separate from the corticospinal tract and are responsible for modulating the movements generated through that primary pathway frontal eye fields area of the prefrontal cortex responsible for moving the eyes to attend to visual stimuli internal capsule segment of the descending motor pathway that passes between the caudate nucleus and the putamen lateral corticospinal tract division of the corticospinal pathway that travels through the lateral column of the spinal cord and controls appendicular musculature through the lateral motor neurons in the ventral (anterior) horn lumbar enlargement region of the ventral (anterior) horn of the spinal cord that has a larger population of motor neurons for the greater number of muscles of the lower limb premotor cortex cortical area anterior to the primary motor cortex that is responsible for planning movements pyramidal decussation location at which corticospinal tract fibers cross the midline and segregate into the anterior and lateral divisions of the pathway pyramids segment of the descending motor pathway that travels in the anterior position of the medulla red nucleus midbrain nucleus that sends corrective commands to the spinal cord along the rubrospinal tract, based on disparity between an original command and the sensory feedback from movement reticulospinal tract extrapyramidal connections between the brain stem and spinal cord that modulate movement, contribute to posture, and regulate muscle tone rubrospinal tract descending motor control pathway, originating in the red nucleus, that mediates control of the limbs on the basis of cerebellar processing stretch reflex response to activation of the muscle spindle stretch receptor that causes contraction of the muscle to maintain a constant length supplemental motor area cortical area anterior to the primary motor cortex that is responsible for planning movements tectospinal tract extrapyramidal connections between the superior colliculus and spinal cord vestibulospinal tract extrapyramidal connections between the vestibular nuclei in the brain stem and spinal cord that modulate movement and contribute to balance on the basis of the sense of equilibrium working memory function of the prefrontal cortex to maintain a representation of information that is not in the immediate environment
This work, Anatomy & Physiology, is adapted from Anatomy & Physiology by OpenStax, licensed under CC BY. This edition, with revised content and artwork, is licensed under CC BY-SA except where otherwise noted.
Images, from Anatomy & Physiology by OpenStax, are licensed under CC BY except where otherwise noted.